
One of the crucial tools of modern evolutionary thinking is the notion that it is not only necessary to think about how a particular trait or relationship might be beneficial to its holder now, but also what circumstances were required for it to have become an adaptive variation in the first place. It is also essential to consider what structural factors must remain present to keep a trait from being removed from a population – is the behavior an evolutionarily stable strategy, or one which is subject to invasion by more “exploitive” traits? In this context, then, what are the biological-fitness enhancing values of “proto-culture” or communication which might have encouraged its emergence in early humans? Clearly this question is closely related to, and dependent upon, the adaptive value of cognition, consciousness, and communication. Is the continued existence of cultural behaviors which seem biologically maladaptive simply an unavoidable consequence of having big brains and jabbering mouths? Or are there group selection benefits? Are cultural behaviors subject to the same constraints of biological fitness, or do they reside in some other selective regime?
It might be useful to distinguish between discussions of (1) the adaptiveness and history of the evolution of culture and information transmission, (2) how evolutionary forces might act to generate or explain specific instances of culture, (3) social- or culturally-based forces which might drive dynamic “evolution” of culture, and (4) the effects of cultural transmission on biological evolution. Although they are all interrelated, and hopefully addressable by comprehensive models, they are often treated as distant fields that happen to have overlapping terminology. Some anthropological or historically oriented perspectives work to explain the events and dynamics of culture in traditional human or psychological terms but seem to lack the ability to generalize or make strong connections to other fields. Radical meme-theorists often extend Richard Dawkin’s concept of an informational replicator to the point where all human behavior is controlled by self-replicating concepts, all trying to out-reproduce each other by spreading to as many individuals as possible. This stands in stark contrast to the conservative biological approach which suggests that any influences of cultural transmission are slight, and must always be subservient to rules of hard-coded biologically adaptive behavior. I’m interested in perspectives which are integrative – meaning that they are extensible and explanatory on multiple scales. Especially attractive are models which are capable of coping with the very high speed (orders of magnitude more rapid than generation turnover) changes in “superficial” cultural traits and information, as well as providing insight about longer term trends.
The tricky part is that to evaluate any hypothesis for the adaptive value of culture, a model with very specific mathematical properties must be defined. There are many plausible models, each implying a different adaptive argument and possessing different dynamical properties. Distinguishing among the models will require a great deal of empirical data, most of which have yet to be collected. And there is probably a limit to how far it is worth pushing the models – the intent should be more to explain patterns than specific instances.
We can easily explain by a sociobiological hypothesis why humans have symbolic capacities for culture. The main adaptive functions include interpersonal communication and memory organization. Both of these functions contribute to more efficient social learning and hence to more effective use of the various advantages of cultural transmission. However, since these functions are served equally well by any well organized symbolic system, we cannot explain much of the difference between the structures of different cultural systems by adaptive arguments. Language is the prototypical case: Chinese is not an adaptation to life on the Yellow River Plain, nor is English an adaptation to living in the British Isles, except in the very limited sense that its is useful to speak the language that happens to be common in a given place. (Boyd & Richerson 1985, p. 274)
Robert Boyd and Peter Richerson have done some work on integrating mathematical descriptions of cultural and biological transmission. They define culture “…as the information affecting phenotype acquired by individuals by imitation or teaching” – although they mostly restrict their analysis to a consideration of effects on the time scale of generations. Their book provides quite a few adaptive explanations for culture.
In evolving a reliance on cultural transmission the human species may well have ‘traded’ high rates of random error caused by individual learning in variable environments for a lower rate of systemic error (with respect to genetic fitness) due to the partial autonomy of cultural evolution. (Boyd & Richerson 1985, p. 289)
Essentially, the argument is that if individual experimentation is “costly”, it might be better to just adopt the behavior of successful group members. If the environment is sufficiently variable in time or patchy in space, it might be beneficial for groups to be able to adapt or adopt new behaviors more quickly than biological evolution would easily allow. In addition to allowing rapid change, the plasticity of cultural evolution may act to increase the diversity – the raw material for adaptive change – of strategies which are present in a population. In a set of computer simulations of evolutionary games, Findlay, Hansell and Lumsden (1988) showed that in their model “…biocultural games show a greater diversity of dynamical behaviors than their purely biological counterparts, including multiple fully polymorphic equilibria…biocultural games on average show greater equilibrium strategy diversity. These results suggest that cultural transmission in the presence of natural selection may be an important mechanism maintaining behavioral diversity in natural populations” (p. 245). To put it another way, models which allowed strategies to be transmitted both culturally and genetically displayed more attractors – bio-culturally stable strategy mixtures for the population to explore, than models of biological evolution alone.
It is also important to realize that for social creatures like humans a large part of the environment which determines their fitness consists of other humans. Survival may at times depend on an ability to coordinate with others for various kinds of collective action. So the adaptive environment in which cultural traits are “evaluated” will consist partially of the traits of other individuals. Findlay, Hansell and Lumsden (1988) refer to the concept of bio-cultural fitness, using it as a framework for thinking about the combined effects of genes and culture on the fitness of a behavioral phenotype. In organisms that have culture, the two types of fitness may be coupled because once cultural systems have evolved, the potential biological impacts of failing to adopt local cultural traits could be large. It is easy to think of situations where an individual’s ability to procure resources or a mate might be impaired if they speak a different language or follow a different system of courtship. “Cultural traits which affect mating preference could similarly affect genetic evolution through the action of sexual selection.” In a sense, “the human genome could be ‘domesticated’ by culturally transmitted traits” (Boyd & Richerson, 1985, p. 277). These effects may not be unique to humans. A study of mate choice copying in guppies gives “…strong evidence for the role of non-genetic factors in sexual selection and underlines the need for new models of sexual selection that explicitly incorporate both genetic and cultural aspects of mate choice” (Dugatkin & Godin 1992, p. 179).
As I began to suggest in the previous section, the possession of cultural properties might make it necessary to revitalize some limited group selection arguments. Culture makes possible a certain degree of “fitness sharing” across individuals in a group and, as much as groups are homogeneous in their cultural traits, some degree of competition between genetically equivalent groups. Imagine, for example, the relations between groups which emphasize “pacifistic” and “warlike” ideals. Again, the outcomes can be highly complex and unintutive (there may be many situations where a collectivist pacifist society without “mean” neighbors does the best), and it is important to be highly cautions about drawing sociological conclusions. My purpose is simply to point out reasons that the inclusion of cultural transmission and other “humanistic” information effects is both necessary and helpful. Humans also possess many other properties which are interesting from an information perspective and may be pre- or co-requisites of cultural adaptation: strong language, abstract reasoning, symbolic skills, deception, intentional communication, etc.
… During hominid evolution, information could have been transmitted through leisure-time social interaction, vocal communication, material culture such as tools and art, and later, speech. Thus although differences in active information transfer, like the ones involved in symbolic communication, are undoubtedly quantitative, a truly significant product of human evolution is the ability and tendency to engage in active information transfer of all kinds. (King, 1991, p. 98)
Models expressing the properties of cultural transmission in humans must be capable of allowing for effects of intentionality and rationality in action and communication. That is not to say that people are always (or even frequently) rational in their behavior, but that they are capable of figuring out the “rules of the game” and exploiting them to their own ends. In many cases this does not actually conflict with adaptive arguments: over time the processes of evolution can be nearly as creative about testing limits as individuals are, so strategies which are evolutionarily stable are usually not susceptible to “cheaters.” However, individuals are capable of making decisions (and convincing others to abide by them) which are not easily explained with a simplistic adaptive argument – becoming a celibate monastic hermit for example, or fighting and dying for an abstract cause.
In a very real sense, all of these models and hypotheses, no matter how reasonable or well thought out, are “just so stories.” Each has a set of sometimes hidden assumptions which must be justified before the predictions will hold true. One assumption which, although it is not unreasonable, is a requirement of many models of cultural change, is that traits are discrete. They do not always need to be discrete in the sense of having only integer values, but in the sense that when an individual is exposed to trait A and a variant A’, one or the other is adopted rather than a combination of both. This is related to the property of heritability which is a requirement for evolutionary processes. Most of the models which are based on blending inheritance lose the capability for the gradual incremental drift which is the essential raw material for evolution by selective processes. This brings forward again the question of the scale and nature of the “fundamental units” of culture and information. This is a recurring theme – along with the discrete vs. continuous trait debate. Part of the reason it is so difficult to apply evolutionary logic to social and cultural phenomena is that it simply isn’t clear what the nature of the substrate is – how similar the properties are to those of DNA substrate for encoding genes. Even when solid numbers on sociological variables can be obtained, the discrete nature of the data can be an artifact of the measurement instrument. For example, voting records and political party affiliation could be treated as an indicator of political alignment, but they are probably discretized measurements of the continuous and dynamic collection of variables which describe an individual’s political perspective.
| Fig. 3 Graphs indicate theoretical probability of a “naive” individual acquiring a cultural variant as a function of the frequency of the variant among the individuals potential models, assuming: (a) unbiased transmission, (b) directly biased transmission, and (c) frequency dependent biased transmission. (copied without permission form Boyd & Richerson 1985, p. 207) | 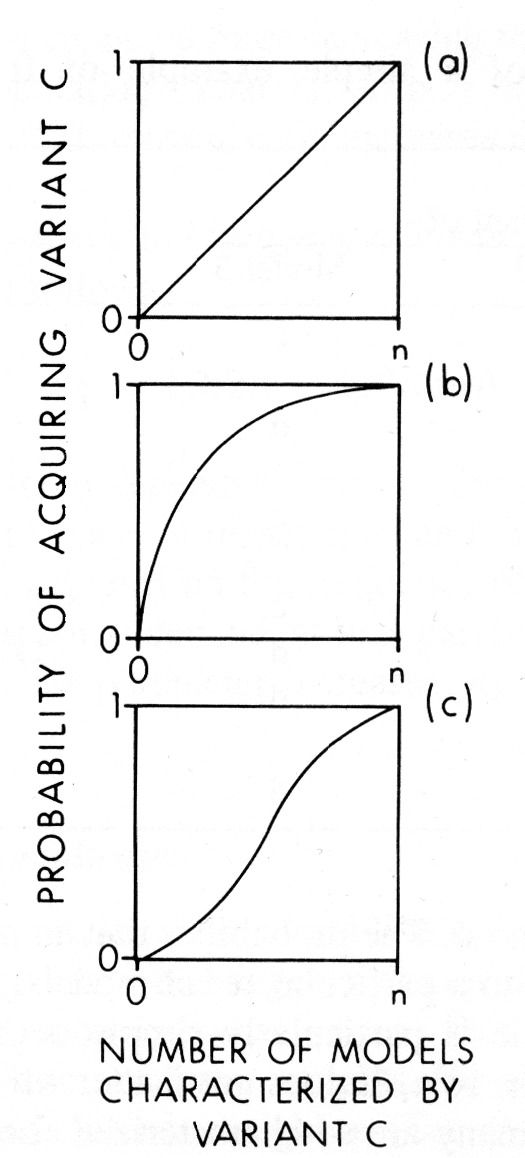 |
The model Boyd and Richerson (1985) develop is presented in several steps. They first consider the effects of “vertical” transmission, or adoption of traits displayed by biological parents. This is strongly analogous to biological transmission. They then consider “oblique” transmission – adoption of traits displayed by other mature models of the parents’ generation. And then they briefly consider “horizontal” or peer-peer adoption. As I already mentioned, most of their models consider relatively slow moving heritable traits like language or religious affiliation. I have not examined the models of horizontal transmission in enough detail to know whether they could describe the fast transmission necessary for a description of gossip or institutional structure. The results of the analytical solutions of Boyd and Richerson’s models of various bias effects on traits are fascinating. In this context, bias refers to systematic misperception or filtering effects triggered by some kind of perceptual cue. This will be discussed further in the section on human communicative processes. Bias is a useful concept because its effects do not need to be deterministic. A bias is simply a predictable tendency, a nudge in a particular direction. There is plenty of room for noise and free-choice in a bias model, as the idea is that although the effects are due to processes occurring in each individual, they may only be detectable as a trend in a population or aggregate measure – a deviation from what might have been expected.
Boyd and Richerson describe three main bias effects which would impact the expected dynamics of cultural process (Figure 3):
(1) Direct biases exercised by evaluating the traits to be adopted by their own properties.
(2) Frequency dependent bias in which commonness or rarity of traits is used as the criteria for adoption.
(3) Indirect biases which result from choosing models based on index traits, after which other traits are imitated without further evaluation. (p. 284)
Indirect biases especially allowed for strong “peacock’s tail” effects – situations in which biological selection feedback causes arbitrary traits which are originally used as indicators of fitness in sexual selection to “run away” to values which are sometimes ridiculously non adaptive. “If we are correct, the forces of guided variation and direct bias are often weak enough to lead to substantial cultural variation, and therefore [biological] natural selection will be an important force acting to change the frequency of different cultural variants” (Boyd & Richerson, 1985, p. 285).
It is frequently the case that arguments from biological natural selection are used to explain changes at the cultural level. But, despite the fact that I’ve spent a great deal of space discussing the potential impacts of culture on biology, biological fitness may not be the best place to look for explanations cultural changes. It is possible that in some instances cultural and genetic transmission are largely decoupled. The current properties of culture may be such that many of the traits are effectively neutral with respect to biological fitness or are able to overpower the bio-fitness constraints because of runaway processes like indirect bias. In this scenario the cultural fitness of a trait need no longer be related to the lifetime fitness of the individuals possessing it. In fact, the reproductive fitness would seem to be a poor indicator of cultural fitness for fast moving traits. In other words, it may be possible for culture, especially symbolic culture which is already fairly arbitrary, to evolve and follow a selective regime which is not necessarily congruent with the biological one. Traits could have a fitness in the sense of differential reproduction, measured by how many individuals adopt them in successive points in time. Unknown parameters could be acting to drive selection for various cultural or informational traits in the same way in which selective functions applied to the computerized worlds of genetic algorithms8 result in “evolutionary” change. The strongest version of this would be meme based theory, but the principle can still hold in the more palatable sense of human biologically influenced intentionality and unconscious choice, developed over evolutionary time, as the selective force on cultural properties. In other words, there could be biological selection for various cognitive structures which then become the selective “environment” in which cultural properties are tested.
To me, a useful way to view the relationship between cultural and biological selection is with the metaphor of co-evolution. In the same way that two interdependent species act to deform each other’s fitness landscape through an ongoing arms-race of co-evolved traits, biological and cultural phenotypes could exist in different selective regimes, but still be coupled to each other through each individual’s actions so that all of its behavior is affected by both regimes. A phenotypic trait can have values in both the cultural and biological domains. Some behaviors or traits will be selected for in one regime and be neutral in the other. In some cases culture may reinforce the biological selective pressures, and in others conflict with it, the resolution eventually occuring through drift in other trait dimensions.
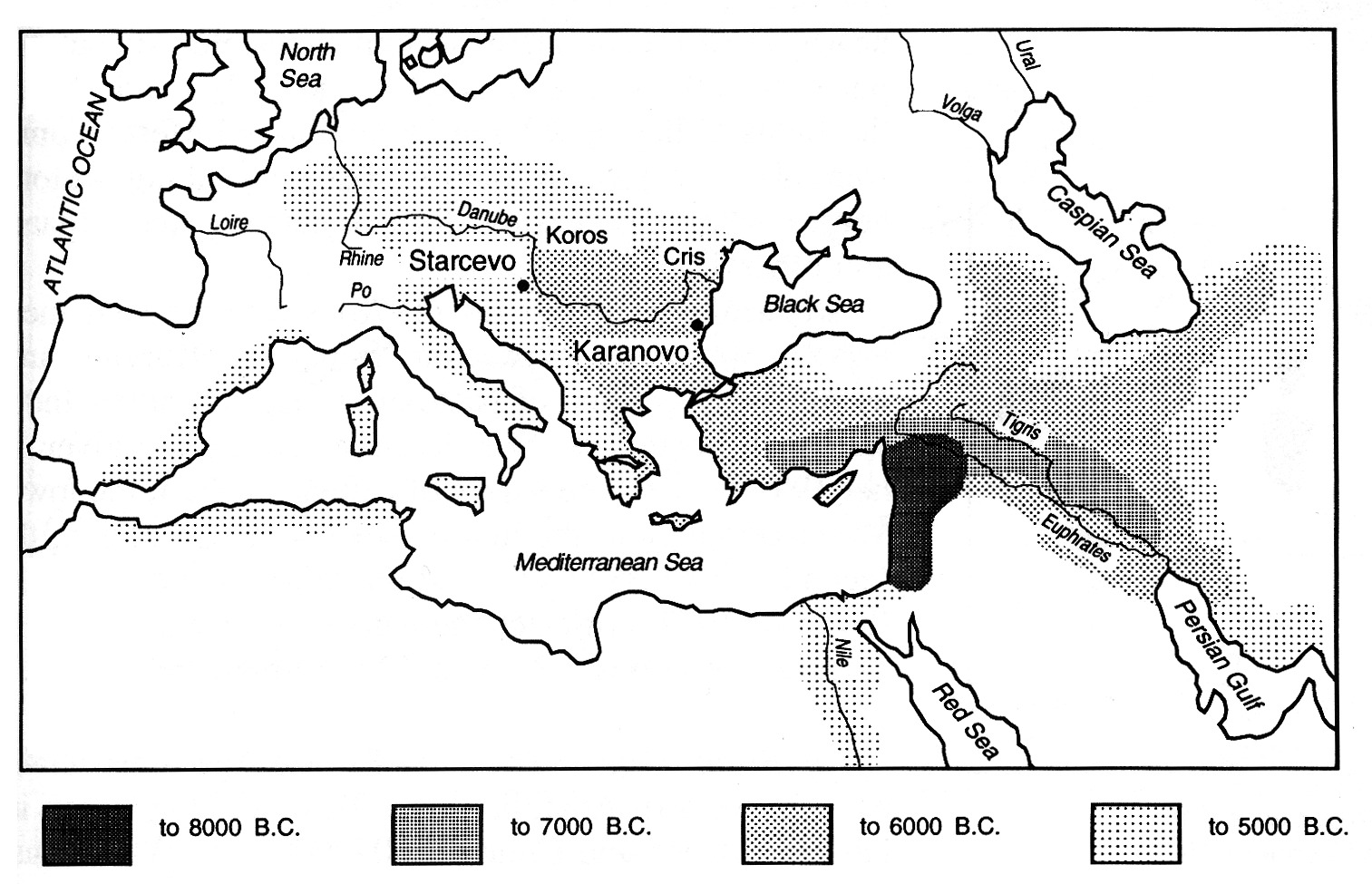 |
| Fig. 4 Map of the spread of Neolithic farming cultures from the Fertile Crescent. Shading indicates presence of archeological indicators associated with agriculture for given time period. (copied without permission from Cavalli-Sforza, Menozzi, and Piazza 1994 p. 257) |
One model for which the discrete/continuous distinction may be less important is that of culture as a process of innovation diffusion. The idea is that an innovation or idea occurs in one location and diffuses spatially through a population in time as if it were a biological epidemic or a chemical dye in a medium (as in the Blue Tit example). The concept is useful in that it allows aggregation of all the variables of individual choice and action so that the process of transmission can be viewed as a probabilistic “random walk” or percolation of a trait through the population – an area of mathematics which is fairly well understood. Some, especially historical processes, appear to be well described by such a model, although diffusionist perspectives seem to be currently out of favor in anthropology. The archeological record seems to show Neolithic agricultural practices spreading through Europe in such a fashion. (Cavalli-Sforza, Menozzi, & Piazza, 1994, See Figure 4) Although the diffusion concept may be a good metaphor on some (very large) length and time scales, it seems to lack many of the qualitative features of culture as it is currently experienced. Certainly there are very strong spatial effects on some traits, (language, regional dialects, accents, etc.), but as contacts between people become increasingly de-spatialized through the use of travel and electronic media, it becomes difficult to imagine the dimensions on which diffusion could be observed. Furthermore, diffusion concepts would seem to ignore the potentially powerful effects of social structure: long distance contacts, local friendships, ghettoization, political affiliation, mass media, etc.
A solution might be to create a model capable of including the effects of structural biases introduced by the effects of social structure, while still allowing probabilistic descriptions and limited diffusion. To me an exciting possibility would be the integration of diffusion models with social network descriptions of contact between individuals and contact with their environment. Trait or information diffusion could be imagined as percolation along the ties between social actors with probabilities weighted by the frequency of contact between them. This would have the advantage of including Boyd and Richerson’s (1988) concepts of vertical, horizontal, and oblique transmission in the more general description. In other words, transmission of traits from parents to offspring could be due to the strong tie (extremely high contact probabilities) between parents and their offspring during childhood – yet long-distance, peer-to-peer, and media effects could be included as well. This idea will be discussed further in the section on social networks.
I think that it is important to reemphasize the role of material culture in the transmission of information and cultural traits. Human-generated artifacts have the ability to carry information and should properly be considered alongside direct personal communication. This was alluded to in the quote from King. Art, architecture, artifacts, tools, weapons, and more recently, texts, recordings, and photos all have strong communicative potential and have a remarkable ability to cross space and time. Material resources and positions in social structure (wealth, class, education level) are often heritable and transmissible (Bowles & Gintis, 2001, Boyd & Richerson, 1985). And social structure will likely influence contact with the information embedded in material culture as well as the information carried by individuals. This means that artifact- or media-based transmission can also be described from a percolation-network perspective. This could be done either directly with descriptions of specific contact, or by considering the correlated environment effects which might appear because of social network associations with individuals. In other words, increased contact with a friend will probably mean increased contact with the friend’s media and material culture as well – both via second-hand effects and simple proximity. For example, I’m much more likely to attend a show with a theater-going friend, read the magazines in his bathroom, etc. One advantage of this perspective is that it essentially sidesteps the correlated environments problem of previous attempts at separating the developmental effects of genes and culture in hereditary behavior transmission – the environment is the culture. Clearly it is important to consider what the term information “means” in these contexts. Can its properties be described in a sense which is discrete and/or relevant to questions about social and material culture transmission?
Very interesting research. I am concerned with the dissimentation of information amongst cultural groups via traditional media such as oral transmission, gossip etc.