
“Culture … is that complex whole which includes knowledge, belief, art, morals, law custom, and any other capabilities and habits acquired by man as a member of society.”
-Sir Edward Burnett Tylor (Primitive Culture, 1871)
One of the things which initially prompted me to do research on information transmission and culture was the question of the relationship between genes and culture. I was in an evolution class and doing reading on dynamical systems at the same time. So the question wasn’t so much the near cliché of Nature vs. Nurture, but more how the cultural and genetic influences might play out as a developmental system for generating an individual’s phenotype. There seems to be an odd asymmetry in the evolutionary literature. The mathematical framework for describing the changes in population gene frequencies brought about by differential selection on organisms is very well developed. Many of the implications of the powerful concepts of genetic evolution and fitness lead to explanations of certain social phenomena which are both fascinating and persuasive (kin selection and altruism, for example). But it seems that in many evolutionary models surprisingly little attention is paid to the potential impact of cultural transmission in determining some aspects of the “behavioral phenotype.”
There are many reasons for this, I’m sure. We now understand a great deal about the actual physical process of genetic transmission, and this makes it possible to calculate the statistical mechanics of genetics with some certainty. Although there has been a long tradition of cultural research in anthropology and the other social sciences, the models and metaphors employed do not always translate easily into mathematical terminology (and vice versa). Despite attempts at interesting “meme” analogies, and some philosophical musings on the nature of replicating information in general2, there is no understanding of what the fundamental units of culture might be. The challenge is to create descriptions of cultural process which are in a form which allows comparison and integration with genetic descriptions. The specific adaptive values of culture are poorly understood, and the fitness implications of bio-cultural transmission are only beginning to be examined. At this point humans are the only species which certainly include some amount of cultural transmission processes in the construction of behavioral phenotypes3. The uncertainty is due in part to considerable uncertainty about how culture should be defined in humans, let alone in a form general enough that it could be tested for in other organisms. But intuition, and some evidence, suggests that forms of information and behavioral transmission do exist in groups of other animals. Examination of these potential “proto-cultures” may help define the underlying ideas of culture and information transmission.
The study of animal behavior and learning is an old and extremely active research area. I cannot pretend to have more than a passing familiarity with the literature, but it seems possible that some of the phenomena of information transmission might not even require a clear ability to learn new behavior at the level of the individual. An alarm reaction in a school or herd, for example, could be an entirely programmed response, where the activation of the response might employ a crude form of social communication. A single individual may notice the presence of a predator and communicate this environmental discovery to others through a behavioral display. This behavior activation might spread through a group in an epidemic fashion, perhaps causing a mass evasive action in which all individuals, including those who never directly sensed the predator, will participate. (Smith, 1991) Clearly, information about the state of the world is being transmitted, even without the requirement of a sense of intentionality of the participants. Boehm (cited in King, 1991, p.100) presents a “…hierarchy of communication intentionality” “… suggest[ing] that an animal might: (1) advertise emotional states only; (2) communicate information about its social and physical environments; (3) intend to communicate so that the information transmitted has an effect upon the hearer; or (4) communicate intentionally in matters that involve displacement in time or space.”
But how should the role of individual learning be viewed in the process of cultural transmission? Organisms may live in a group, but if individuals are able to learn to make distinctions about environmental states without the assistance of interaction with other members, cultural transmission may not be required for the acquisition of behavior. Still, the presence of conspecifics may help facilitate the process through socially mediated learning. Laland and Williams (1997) describe a series of laboratory experiments in which schools of fish are trained to locate a food source. When “naive” fish are introduced, they follow the school of demonstrators to the food. The location of the food is “remembered” by the fish (or perhaps the school) even when all of the original fish have been replaced. “The results suggest that the tendency to shoal may facilitate a simple form of guided social learning, which allows guppies to learn about their local environments. They also imply that selectively neutral behavioral alternatives may be maintained as traditions in aggregated animal populations by very simple social mechanisms.” (p. 1161) This is a case which certainly doesn’t require any intentionality, and in which there is no abstract communicative process; yet it is consistent with some definitions of transmission: “…social transmission … occurs when a social interaction increases the likelihood that one individual will exhibit a behavior initially the repertoire of another.” (King 1991, p. 99, from Galef)
I think that the guppy example emphasizes the need to distinguish between the transmission of information and the transmission of culture. I will save a discussion of the nature of information for later, but I think that Galef’s statement provides a good working definition of low-level social information transmission. A good definition of cultural transmission might require the inclusion of a clear description of the kinds of learning and the time frame to be considered. I’m going to define cultural transmission as occurring when a behavior initially in the repertoire of one individual appears in the repertoire of another, but only when the second individual also gains the ability to re-display the behavior at points in time and space which are appropriate but distinct from the original setting. For a setting to be considered “appropriate,” it would probably have to preserve an association with some of the perceptual and contextual elements of the initial situation. Two further examples point out some of the complexities in trying to arrive at any kind of working definition of culture or cultural transmission.
MIT’s StarLogo4 website presents a model which simulates the synchronization of firefly flashing. The model is a translation into computer code of a simplified description of the flashing behavior of one firefly, and a set of rules for how each of the single firefly “agents” will interact with its neighbors. Although it is not clear that the model presented in the simulation reflects the actual process as it occurs in real fireflies, the dynamics of the simulated flies are qualitatively similar to what is observed in nature. (Buck 1988)
In the region from India east to the Philippines and New Guinea, enormous aggregations of fireflies gather in trees and flash in near-perfect synchrony. While different species have slightly different methods for flashing in rhythm, the behavior that is modeled here is governed by the following rules:
– Each firefly has an intrinsic flashing frequency, and when left alone it will flash at periodic intervals.
– The flashes are timed by the progressive excitation of a chemical within each firefly; the excitation increases until it reaches a certain threshold, at which point a flash is emitted and the excitation is reset to zero.
– If a firefly senses a certain amount of luminescence from its neighbors, it will reset its excitation to zero in order to flash simultaneously with those neighbors in the future; however, if the excitation is close enough to the flashing threshold, the flash has already been started and will proceed as planned even though the excitation is reset to zero. (The signal to flash is initiated when the excitation is 80% of the way to the threshold value.)
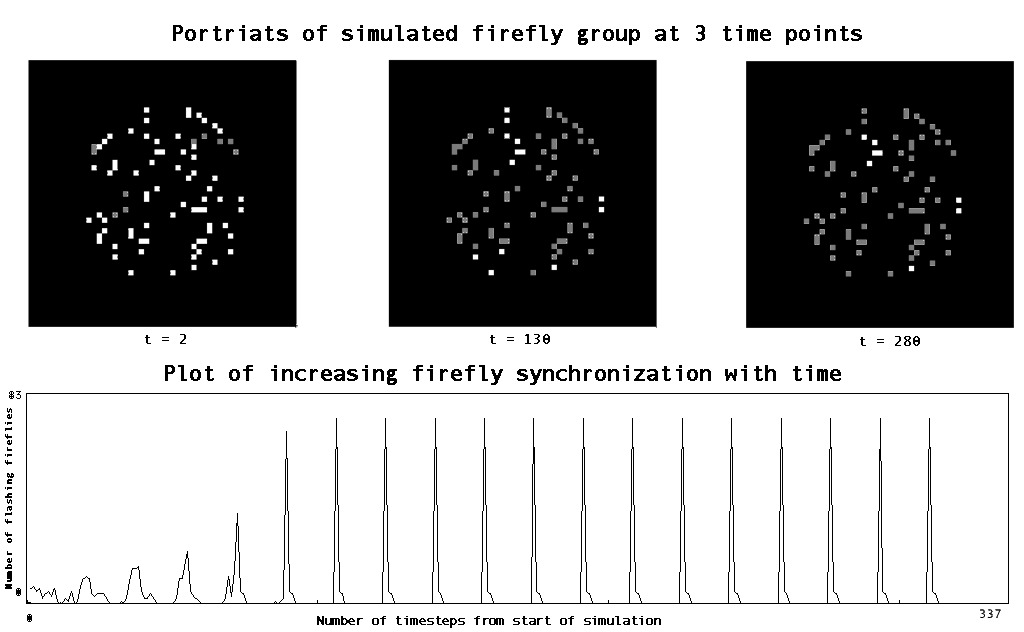 |
| Fig. 2 Screen shots and time series from StarLogo simulation of firefly flashing behavior. In the top panels, gray squares indicate flashing fireflies, white squares dormant fireflies. Time series shows the number of fireflies “on” at each time step. Notice the increasing synchronization – development of large spikes with almost no tails. (code from StarLogo web site 2001, slight modifications by author to produce time series.) |
Obviously real fireflies do many more interesting and complex things than this model allows. But the important point is that the model demonstrates concretely the formal potential for coordination – the emergence of a specific form of synchronized group signaling – among a large number of individual “agents” relying only on very simple rules and with no central coordinating authority. The only form of communication taking place among the simulated agents is a simple on/off message which can only be received by the few agents in the immediate vicinity. This a good example of the type of phenomenon investigated by complexity theory. The synchronized flashing of the group is considered an “emergent” behavior because, although it is definitely a result of the interaction rules given to the agents, it is not explicitly described in the rules. Furthermore, it is a property of the system that is scale-dependent – it doesn’t have any meaning and could not be discovered by examining the behavior of one firefly in isolation. Although it would be quite a stretch to class the firefly example as a cultural phenomenon, there is clear evidence of information transmission, (increasing display of identical flashing frequency) and even of a type of aggregation or consensus formation. The consensus achieved in this model is qualitatively unlike “issue” consensus in humans, because the message is discrete and the range of transmission increases as each of the sub-groups synchronize into bright regions. But it is interesting to think about comparing the firefly model to some instance of human behavior – synchronized clapping behavior in concert crowds for example. (Fountain, 2000)
Another classic case concerns the reports of British birds opening milk bottles. Apparently, sometime in the late ’40s, Blue tits began pecking through the foil tops of milk bottles to drink the cream. In his brief discussion, Bonner (1980) implies that the innovation appeared in a single location and the skill spread all over the British Isles through a process of epidemic imitation and cultural diffusion. However, even the paper he cites (Hind & Fisher, 1951) offers a more cautious interpretation: the apparent cultural transmission may be better described as a socially facilitated learning process of “local enhancement.” Although the distinction may seem fairly arbitrary, the implications are interesting. If the birds learn the behavior though a local stimulus enhancement phenomenon, there is much greater potential for birds to make the discovery independently. “Sherry and Galef’s results, for example, showed that experience with previously opened bottles was sufficient to establish milk bottle opening in birds that do not spontaneously exhibit the behavior” (Nicol, 1995, p 83)
In a sense, in the case of the Blue Tits the distinction between imitation and stimulus enhancement learning is not important – behavior is still moving from one repertoire to another, and the behavior still spreads whether from one genius bird or several. But clearly there may be situations where the distinction is essential – when birds are kidnapped to a behavioral laboratory and must learn arbitrary new tasks to obtain food, for example. Nicol gives a good description of imitation:
… the observer may be able to copy the exact motor patterns of the demonstrator by some (as yet unknown) process of cross-modal matching.
as distinct from stimulus enhancement:
… the behavior of the demonstrator may result in an increase in the salience of a particular stimulus or location within an overall environment for the observer. The observer’s attention may be drawn towards previously irrelevant features, or the observer’s motivation to investigate the stimulation or location may be increased. (Nicol, 1995, p. 83)
One way to think about this is to imagine what kind of meaning content would need to be conveyed in each of the suggested possibilities. In the case of socially mediated stimulus enhancement, there is a sense of information as discrete incidence, a releaser of innate action: “this bright object is food, exhibit appropriate behavior.” (The firefly and alarm-reaction examples would fit into this class.) Imitation would seem to require the transmission and reception of information of a more narrative or descriptive kind: “this is a sequence of behaviors which you can use to get food from an unusual source.” It is not clear that either process would be in any sense less complex than the other. And, as Nicol points out, “… stimulus enhancement followed by trial and error learning may be a more efficient way of acquiring a skill in many circumstances.” (p. 84) Part of what is confusing here is that when considering “lower” animals, imitation would seem to be the more complex task – perhaps requiring the abstract concept of the Other as similar to the Self.5 In humans, where the ability to abstract is fairly strong, intentional communication via stimulus enhancement is arguably one of the highest forms of communicative cultural behavior, and imitation is viewed as a relatively mindless task.6 I believe that the root of the confusion lies with the automatic attribution of intentionality and the capability for abstraction which we, as humans, naturally give to descriptions of other humans.
The distinction between the processes of transmission-by -stimulus-enhancement and transmission-by-imitation becomes crucial if we are concerned with what happens when errors, mistakes, or misinterpretations occur between the communicators. The results of a chain of transmission events could be very different if the message is singular as opposed to being constructed from continuous or multi-part items. In the multi-part case there is the potential for behavior patterns to “drift” – errors can be additive in a way which is impossible in the singular case. One way of thinking about this is to refer again to the Blue Tit example. There are several ways of describing the communicative process which occurs when one bird observes another opening a bottle and obtaining food. Imagine that the message received by the observer is a singular one: it only needs to reclassify the milk bottle from “objects which are not food” to “objects which are food” The observer must rely on previously learned or genetically programmed behavior to determine the actions appropriate for getting food from a bottle. Errors in the message might result in the bird attempting to mate with the bottle, but if the bird tries to get food, it will always utilize the same patterns of behavior. If the patterns are genetically determined, they may be fairly uniform across a population, and changes must take place over evolutionary time through selection.
If the process is one of imitation and learning in which a complex series of motor skills must be correctly perceived and emulated, there are many opportunities for errors to disrupt the acquisition of the behavior so that it cannot be successfully utilized. However, there is also the possibility that a misperception on the part of the observer will result in a behavior which is more effective in obtaining food than the one he observed – perhaps using claws rather than beak to remove the foil. In other words, if the message consists of a collection of units or a continuous variable rather than a single programmed stimulus, there is the potential for errors or individual innovation to create many differing versions of a behavior. The stability of a predetermined behavior pattern is exchanged for flexibility. Although there may certainly be interesting fitness implications (in the standard biological sense) for birds which have different variants of a particular transmitted or learned behavior, at the moment I’m mostly concerned with differential selection at the level of the behavior pattern or message itself, rather than on bird reproduction. By allowing errors in learning, the potential may exist for differential selection of behavior patterns on a time scale shorter than an individual’s life span.
I’m aware that it is a questionable jump to begin applying selectionist terminology to a cultural process, but I believe that in this case it is justified. That is, we could make the not-too-unreasonable assumption that birds who aren’t able to open bottles will stop attempting to use unsuccessful behavior. Alternatively, if the observers do not “see” a reasonable reward obtained, they may not try to copy the bottle-attacking behavior of the demonstrator birds. Or in the strongest case, they may evaluate the effectiveness of their neighbor’s strategies and adopt the best. Either way, some variations of the behavior might increase in frequency in the population at the expense of others. This hypothetical example has many problems 7, and would of course be highly behavior-, animal-, and environment-specific. Over time, differential transmission of behavioral techniques could conceivably lead to the discovery and diffusion of very effective and/or very locally adaptive behaviors in a population. If there was any degree of restriction of the “mixing” or diffusion of traits through the population, subgroups which employ different behaviors to accomplish the same task might form. If the behaviors in question were involved with mate choice or courting, there might be effects at the genetic level. Needless to say, the interactions with biological fitness in its classical definition would be complex and difficult to predict intuitively.
Although the cultural components of heredity and behavioral transmission are not as well researched as the genetic component, it is a relatively rich and growing field. I would argue that debates about the nature of the cultural learning and transmission process do not necessarily reduce the usefulness of discussion or qualitative examination in the absence of hard data. But it is essential to remain clear that any conclusions about the dynamics of a message or behavior at a population level will generally be conditional upon the as-yet-uncertain formal properties of the transmission media and message process.
notes
2 Richard Dawkin’s The Selfish Gene (1979) has an extensive discussion of the “meme” concept. Also, see Lumdson and Wilson’s Genes, Mind, and Culture. (1981)
3 This is somewhat of a tautology, as definitions of culture often boil down to “whatever it is that humans do that animals don’t”. But for an extensive listing of papers documenting the existence of vertical (p.50) and horizontal (p.54) cultural transmission in humans, see Boyd and Richerson (1985).
4 StarLogo is an agent-based simulation environment developed for educational purposes. The software and this example can be located at: http://www.media.mit.edu/starlogo/
5 For an excellent discussion and further results of social learning and behavioral transmission in animals, see Zentall and Galef’s Social Learning (1988).
6 It is interesting to contrast the “bulimic” model of education, (cram the students full of facts with the requirement that they spit them out undigested on a test) with the learning process as facilitated by a good teacher (set up the environment so that the conclusion will become evident and let the students figure it out for themselves in their own way).
7 One of the problems is that Blue Tits may not actually imitate – although urban pigeons seem to. See Lefebvre and Palameta (1988).